
در شیمی پلیمر ، پلیمرهای تراکمی به هر نوع پلیمری گفته می شود که فرآیند پلیمریزاسیون آن شامل یک واکنش تراکم می شود (یعنی یک مولکول کوچک مانند آب یا متانول به عنوان محصول جانبی تولید می شود). پلیمرهای چگالشی با چند تراکم تشکیل میشوند، زمانی که پلیمر با واکنشهای تراکم بین گونهها با تمام درجات پلیمریزاسیون ، یا با پلیمریزاسیون زنجیرهای متراکم ، زمانی که پلیمر با افزودن متوالی مونومرها به یک مکان فعال در یک واکنش زنجیرهای تشکیل میشود، تشکیل میشود . اشکال جایگزین اصلی پلیمریزاسیون پلیمریزاسیون زنجیره ای و پلیمریزاسیون هستند که هر دو پلیمرهای افزودنی را ایجاد می کنند .
چند متراکم شدن پلیمریزاسیونی که در آن رشد زنجیره های پلیمری با واکنش های تراکم بین مولکول های تمام درجات پلیمریزاسیون انجام می شود. یادداشت: 1. مراحل رشد به صورت زیر بیان می شود: P x + P y → P x+y + L {x}∈{1,2,…∞};{y}∈{1,2,…∞}
که در آن Px . و Py به ترتیب نشان دهنده زنجیره های درجات پلیمریزاسیون x و y و L یک محصول جانبی با جرم مولی کم است 2. اصطلاح قبلی «چگالش چندگانه» مترادف با «پلیمریزاسیون تراکمی» بود. تعاریف کنونی پلیمریزاسیون و پلیمریزاسیون زنجیره چگالشی هر دو با اصطلاح قبلی «چگالش چندگانه» پذیرفته شدند.
پلیمریزاسیون چگالی شکلی از پلیمریزاسیون مرحله ای رشد است . پلیمرهای خطی از مونومرهای دو عاملی ، یعنی ترکیباتی با دو گروه انتهایی واکنش پذیر تولید می شوند . پلیمرهای متراکم رایج شامل پلی آمیدها ، پلی استال ها و پروتئین ها هستند . [2] [3]
پلی آمیدها
یک دسته مهم از پلیمرهای تراکمی پلی آمیدها هستند . [4] آنها از واکنش اسید کربوکسیلیک و یک آمین به وجود می آیند. به عنوان مثال می توان به نایلون ها و پروتئین ها اشاره کرد . هنگامی که از اسیدهای آمینه کربوکسیلیک، به عنوان مثال اسیدهای آمینه تهیه می شود، استوکیومتری پلیمریزاسیون شامل تشکیل همزمان آب است:
- n H 2 N-X-CO 2 H → [HN-XC(O)] n + (n-1) H 2 O
هنگامی که از دی آمین ها و اسیدهای دی کربوکسیلیک تهیه می شود ، به عنوان مثال تولید نایلون 66 ، پلیمریزاسیون دو مولکول آب در هر واحد تکرار تولید می کند:
- n H 2 N-X-NH 2 + n HO 2 C-Y-CO 2 H → [HN-X-NHC(O)-YC(O)] n + (2n-1) H 2 O


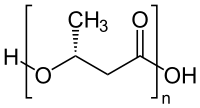
- ساختار شیمیایی عمومی یک نوع پلیمر تراکمی
پلی استرها
دسته مهم دیگر از پلیمرهای تراکمی پلی استرها هستند . [5] آنها از واکنش یک اسید کربوکسیلیک و یک الکل به وجود می آیند. یک مثال پلی اتیلن ترفتالات است :
- n HO-X-OH + n HO 2 C-Y-CO 2 H → [OXO 2 C-YC(O)] n + (2n-1) H 2 O
ملاحظات ایمنی و زیست محیطی
پلیمرهای تراکمی نسبت به پلیمرهای افزودنی بیشتر زیست تخریب پذیر هستند . پیوندهای پپتیدی یا استری بین مونومرها را می توان به ویژه در حضور کاتالیزورها یا آنزیم های باکتریایی هیدرولیز کرد .

چند متراکم شدن
پلیمریزاسیون تراکمی دایمر یا پلیمریزاسیون تراکمی یک فرآیند پلیمریزاسیون ایجاد می کنند است که در آن مواد مختلف واکنش نشان می دهند و یک یا چند مونومر که با یکدیگر متحد می شوند و یک را تشکیل می دهند که در اثر واکنش با مونومرها یا دایمرهای دیگر (یا تریمرها یا تترامرها…) ایجاد می شود. مربوطه افزایش یابد تا پلیمر . روند اولیه بسیار متفاوت از پلیمریزاسیون افزودنی ، که در آن مواد اولیه به نوبه خود به عنوان مونومر عمل می کنند. 1 برای اینکه یک پلی تراکم به طور رضایت بخش انجام شود، مونومرهای تشکیل شده باید حداقل سطح عملکردی 2 داشته باشند، در غیر این صورت زنجیره مونومرهای در حال تشکیل رشد نمی کنند و پلیمر تشکیل نمی شود. در هر اتحاد دو مونومر یک مولکول کوچک از بین می رود، به عنوان مثال: آب. به همین دلیل، جرم مولکولی پلیمر لزوماً مضربی دقیق از جرم مولکولی مونومر نیست.
- 1 ریشه شناسی
- 2 عملکرد
- 3 ویژگی
- 4 طبقه بندی
- 5 همچنین ببینید
- 6 مراجع
ریشه شناسی
اصطلاح polycondensation به محصول جانبی به دست آمده در هنگام پلیمریزاسیون با این روش اشاره دارد، محصول جانبی به دست آمده می تواند آب یا یک الکل کم با وزن مولکولی ، یعنی یک ” تراکم ” باشد.
کارکرد
عملکرد به گروههای عاملی اطلاق میشود که در مونومرها تشکیل میشوند، این گروههای عاملی توانایی مونومر در واکنش با مونومرهای دیگر را تعیین میکنند. سطح عملکرد یک مونومر برابر است با تعداد گروه های عاملی که در مولکول خود دارد. به همین دلیل است که انتخاب موادی که باعث ایجاد مونومرهای مورد استفاده در پلیمریزاسیون می شود، برای دستیابی به حداقل سطح عملکرد مورد نیاز اهمیت حیاتی دارد. برای انجام یک پلی تراکم موفق، در نظر گرفته می شود که حداقل سطح عملکرد باید 2 باشد، یعنی دو گروه عاملی که بخشی از مولکول مونومر هستند.
مثال: HOOC–R 1 –NH 2 (فرمول کلی اسید آمینه، عملکرد 2)
اگر با خودش واکنش نشان دهد، پس:
2 HOOC–R 1 –NH 2 <—-> HOOC–R 1 –NH· + ·OC–R 1 –NH 2 + H 2 O <—-> HOOC–R 1 -NH–CO–R 1 –NH 2 + H 2 O
| عملکرد | |
|---|---|
| 0 | اگر عملکرد مونومرها صفر باشد (هیچ گروه عاملی بخشی از مولکول های مونومر نیست)، ظرفیت آن ها را نخواهند داشت یا واکنش آنها با یکدیگر بسیار دشوار خواهد بود. |
| 1 | عملکرد 1 مونومرها با مونومرهای دیگر واکنش نشان می دهند، اما بلافاصله پس از اینکه عملکرد خود را از دست می دهند، زیرا مونومرها از طریق گروه های عملکردی خود برهم کنش می کنند و متصل می شوند، بنابراین هنگامی که گروه واکنش نشان داد، توانایی انجام دوباره آن را از دست می دهد، و چون تنها یک واحد وجود دارد. گروه عاملی روی مولکول مونومر، دیگر نمی تواند با مونومر یا دایمر دیگری واکنش نشان دهد . |
| 2 | مونومرهای 2 عملکردی قادرند دو بار با مونومرهای دیگر واکنش دهند، که آنها را به کاندیدای عالی برای پلیمریزاسیون تبدیل می کند، زیرا هنگامی که برای اولین بار واکنش نشان می دهند، هنوز یک گروه عاملی دوم را حفظ می کنند که قادر به واکنش مجدد است. این به زنجیره اجازه می دهد تا به طور نامحدود به افزایش جرم مولکولی خود ادامه دهد. |
| ≥3 | عملکرد 3 یا بالاتر به مونومرها اجازه می دهد تا در جهات مختلف رشد کنند، بنابراین آنها می توانند پلیمرهای بسیار شاخه دار ایجاد کنند، حتی قادر به دستیابی به شبکه های ماکرومولکولی سه بعدی هستند. |
ویژگی ها
در صنعت، تمایل به استفاده از موادی وجود دارد که مونومرهای عملکردی 2 را ایجاد می کنند، که گروه های عاملی آنها در انتهای مخالف مولکول یا زنجیره کربنی یافت می شوند، از این طریق پلیمرهای بسیار خطی به دست می آیند. علاوه بر خطی بودن پلیمرهای بهدستآمده، ویژگیهای دیگری که پلیمرهای بهدستآمده از چند تراکم را از پلیمرهای بهدستآمده با افزودن متمایز میکند، برجسته میشوند: 1
- رشد مولکولی کند است و برای به دست آوردن وزن های مولکولی بزرگ به تبدیل مونومر بالایی نیاز دارد.
- همیشه محصولات جانبی با وزن مولکولی کم تشکیل می شود.
- این یک فرآیند گرماگیر است.
دایمر
دایمر (از دو قسمت تشکیل شده است) معانی مختلفی دارد:
- در فیزیک و شیمی، دایمر مولکولی است که از دو واحد مشابه یا مونومر متصل، هر دو توسط پیوندهای کووالانسی و غیرکووالانسی مانند پیوندهای هیدروژنی تشکیل شده است.
- در زیست شناسی، دایمر پروتئینی است که از دو زیر واحد تشکیل شده است. در یک همودایمر دو زیرواحد یکسان هستند و در یک هترودایمر متفاوت هستند.
هترودایمر
هترودایمر به هم است مولکولی که از دو جزء مختلف تشکیل شده است که توسط نوعی پیوند شیمیایی متصل شده اند . نمونه هایی از هترودیمرها هستند پروتئین ها یا پلی پپتیدهایی که از دو زنجیره اسید آمینه با ترکیب متفاوت ساخته شده اند. می گویند در بیوشیمی وقتی یک مولکول از دو جزء تشکیل شده باشد به آن دایمر ، اگر دو جزء یکسان باشند یک همودایمر تشکیل می دهند، در حالی که وقتی اجزا با ترکیب متفاوت باشند یک هترودایمر را تشکیل می دهند. 1
هتروتریمر
اصطلاح هتروتریمر به طور کلی برای تعیین یک پروتئین یا پلی پپتید به کار می رود که از سه زیر واحد با ترکیب متفاوت ساخته شده است. 2
هتروتترامر
برای تعیین یک پروتئین یا پلی پپتید استفاده می شود که از چهار واحد با ترکیب متفاوت تشکیل شده است. 2
نمونه هایی از هترودیمرها

- ترانس کریپتاز معکوس ترانس کریپتاز معکوس ویروس نقص ایمنی انسانی است آنزیمی که سنتز DNA از RNA ژنومی ویروسی را کاتالیز می کند و از دو زیر واحد مختلف به نام های p66 و p51 ساخته شده است. 3
- ریلکسین . رلاکسین هورمونی است که از نظر ساختاری یک هترودایمر است که توسط دو زنجیره پپتیدی متشکل از 24 و 29 اسید آمینه که توسط پیوندهای دی سولفیدی به هم مرتبط شده اند تشکیل شده است .
- پروتئین کو . پروتئین Ku موجودات یوکاریوتی است یک هترودایمر که از دو پلی پپتید مختلف (Ku70 و Ku80) تشکیل شده است. عملکرد آن ترمیم DNA آسیب دیده است.
- توبولین . توبولین یک هترودایمر است که توسط دو پلی پپتید ایجاد می شود: توبولین آلفا و بتا. این جزء اصلی سلولی اسکلت میکروتوبول است .
![]()
ساختار کریستالوگرافی ترانس کریپتاز معکوس ویروس نقص ایمنی انسانی این یک هترودایمر است که توسط زیر واحدهای p66 و p51 تشکیل شده است.
یوکاریوتا
| یوکاریوت | ||
|---|---|---|
| محدوده زمانی : 2200–0Ma داشته است. قدیمی، کهنه پروتروزویکو پنکه. پالئوپروتروزوییک – اخیر | ||
 | ||
| طبقه بندی | ||
| دامنه : | یوکاریوتا Chatton 1925، orth. ما عکس 1971 یوکاریا _ مارگولیس و چپمن 2009 | |
| پادشاهی ها و سوپرگروه ها | ||
طبقه بندی تاکسونومیک: 1
در سیستم های دیگر، به جای Protista می رود: 2
طبقه بندی فیلوژنتیک 3 4
| ||
| مترادف | ||
| ||
[ ویرایش داده ها در ویکی داده ] | ||
در زیستشناسی و طبقهبندی ، یوکاریوتا یا یوکاریا (از یونانی : εὖ eu – “خوب”، “خوب”، “درست” – و κάρυον karyon – “مهره”، “سنگ”، “هسته” -) دامنه است ( یا امپراتوری واقعی تشکیل شده اند ) که شامل موجوداتی است که از سلول هایی با هسته های . اسپانیایی سازی مناسب این اصطلاح یوکاریوت یا یوکاریوت است . 5 این موجودات از یک یا چند سلول یوکاریوتی تشکیل شدهاند که از تک سلولی تا موجودات چند سلولی واقعی را در بر میگیرند که در آنها سلولهای مختلف برای کارهای مختلف تخصصی شدهاند و به طور کلی نمیتوانند در انزوا زنده بمانند.
به قلمرو یا امپراتوری یوکاریوتی پادشاهی جانوران قرار دارند ، ، گیاهان و قارچها و همچنین گروههای مختلفی که در پادشاهی پارافیلتیک Protista تعلق دارند . همه آنها شباهت هایی در سطح مولکولی دارند (ساختار لیپیدها ، پروتئین ها و ژنوم )، منشأ مشترکی دارند، و عمدتاً دارای طرح بدنی یوکاریوت ها هستند که بسیار متفاوت از پروکاریوت ها است .
به استثنای برخی از موجودات تک سلولی، چرخه زندگی یوکاریوتی بین فاز هاپلونت و دیپلونت متناوب به دست می آید متناوب می شود که با میوز و لقاح ، فرآیندهایی که به ترتیب سلول های هاپلونت و دیپلونت را تولید می کنند.
- 1 ساختار سلولی
- 1.1 سرجوخه برنامه ریزی کنید
- 2 پخش
- 3 مبدا
- 4 تکامل
- 4.1 تکامل تک سلولی
- 4.2 تکامل چند سلولی
- 5 طبقه بندی و فیلوژنی
- 6 همچنین ببینید
- 7 مراجع
- 8 پیوندهای خارجی
ساختار سلولی

به طور کلی بسیار بزرگتر از سلول های پروکاریوتی هستند سلول های یوکاریوتی و بسیار بیشتر بخش بندی شده اند. آنها دارای طیف گسترده ای از غشاها با هسته احاطه شده توسط پوشش هسته ای ، شبکه آندوپلاسمی و دستگاه گلژی ، و همچنین مکانیسم هایی برای جوانه زدن و همجوشی وزیکول ها ، از جمله اگزوسیتوز و اندوسیتوز هستند . ساختارهای داخلی به نام اندامک مسئول انجام عملکردهای تخصصی در داخل سلول هستند. وجود لیزوزوم ها ، پراکسی زوم ها و میتوکندری ها .
همچنین مشخصه همه یوکاریوت ها یک اسکلت داخلی یا اسکلت درونی است که در این مورد اسکلت سلولی نامیده می شود که از دو شبکه پروتئینی تشکیل شده است: سیستم میکروتوبول و سیستم انقباضی اکتین / میوزین که نقش مهمی در تعیین سازمان و شکل سلولی، درون سلولی دارند. قاچاق (به عنوان مثال، حرکات وزیکول و اندامک)، و تقسیم سلولی. تاژک مشخصه یوکاریوتی و موتورهای مولکولی مرتبط با آن به اسکلت سلولی متصل می شوند.
DNA . قرار دارد سلول های یوکاریوتی در یک هسته سلولی که توسط یک غشای دوگانه نفوذپذیر از بقیه سلول جدا شده است ماده ژنتیکی به چند بلوک خطی به نام کروموزوم تقسیم می شود که در طی تقسیم هسته ای توسط یک دوک میکرولوله ای از هم جدا می شوند . کروموزوم ها تلومرها حاوی هیستون ها ، ، رپلیکون های سانترومرها و . هستند مختلف وجود دارد یک چرخه سلولی مشخصه با جداسازی میتوزی و تولید مثل جنسی توسط میوز . شامل یک مجتمع منافذ هسته ای ، انتقال غشایی RNA ، و الگوهای جدید پردازش و پروتئین در سراسر پوشش هسته ، اینترون ها RNA با استفاده از اسپلایسئوزوم است .
برای مقایسه با ویژگی های پروکاریوتی ، نگاه کنید به: جدول مقایسه .

پلان بدن
سلول یوکاریوتی شکل و توانایی خود را در حرکت تا حد زیادی مدیون اسکلت سلولی است ، زیرا به آن سفتی و انعطافپذیری میدهد. در موجودات تاژکدار، تاژکها را به بقیه سلول متصل میکند و اجازه میدهد تا در حین حرکت یا برای ایجاد جریانهای آبی که غذا را به آن میبرند، ضربه بزنند. در موجودات آمیبوئید گسترش دهند اجازه می دهد تا “پاها” یا شبه پاها را برای حرکت یا تغذیه . همچنین شیارهای تغذیه حفاری ها و کمپلکس آپیکال را اصلاح می کند که به اپی کمپلکس ها اجازه ورود به سلول های انگلی را می دهد.
تنها پس از ایجاد اسکلت سلولی، یوکاریوت اجدادی می تواند فاگوسیتوز را انجام دهد ، زیرا این امر است که از طریق رشد افتراقی فیبرهای خود، باعث تغییر شکل سلول می شود و فاگوسیتوز رخ می دهد. فاگوسیتوز همچنین یکی از ویژگی های اجدادی یوکاریوت ها است، اگرچه در گروه هایی که با روش های دیگر تغذیه سازگار شده اند از بین رفته است. قارچ ها و گیاهان این توانایی را با ایجاد یک دیواره سلولی سفت و سخت در خارج از سلول از دست دادند، اما آنها قبلاً روش های دیگری از تغذیه، ساپروتروفی یا انگلی در قارچ ها و فتوسنتز در گیاهان داشتند.
میتوکندری را که از فاگوسیتوز و همزیستی متعاقب آن یک پروتئوباکتری به دست میآید، به یوکاریوت اجدادی اجازه میدهد تا تنفس هوازی انجام دهد و بنابراین از انرژی موجود در مواد آلی حداکثر استفاده را ببرد. همانطور که در تکامل چنین شخصیت باستانی تعجب آور نیست، در چندین گروه میتوکندری ها آن ظرفیت اجدادی را از دست داده اند و در عوض برای انجام وظایف دیگر اصلاح شده اند. همچنین از یک رویداد درون همزیستی کلروپلاست ها ، در این مورد با یک سیانوباکتری انجام دهند، ، که به گیاهان اجازه می دهد تا فتوسنتز را به وجود می آیند . متعاقباً، گروههای دیگر یوکاریوتها کلروپلاستهای خود را از طریق همزیستی ثانویه با جلبک سبز یا قرمز به دست آوردند.
پخش

علاوه بر تقسیم غیرجنسی سلول ها ( میتوز )، بیشتر یوکاریوت ها دارای فرآیند تولید مثل جنسی مبتنی بر میوز هستند که در بین پروکاریوت ها یافت نمی شود. تولید مثل یوکاریوتی معمولاً شامل وجود یک فاز هاپلوئید است که در آن فقط یک نسخه از هر کروموزوم در سلول ها وجود دارد و یک فاز دیپلوئید که در آن دو وجود دارد. سلول های دیپلوئید از همجوشی هسته ای ( لقاح ) و سلول های هاپلوئید با میوز به وجود می آیند . در موجودات چند سلولی، سه نوع چرخه زندگی متمایز می شود:
- چرخه هاپلونت: ارگانیسم هایی که این چرخه را ارائه می دهند در مرحله بزرگسالی خود هاپلوئید هستند. زیگوت . دیپلوئید است و میوز پس از لقاح رخ می دهد
- هاپلوئیدی را تشکیل می دهند چرخه دیپلونتیک: افراد بالغ دیپلوئید هستند و با میوز گامت های که با هم ترکیب می شوند و یک ارگانیسم دیپلوئید جدید ایجاد می کنند.
- چرخه هاپلودیپلونت : تناوب نسل ها بین افراد هاپلوئید و دیپلوئید اتفاق می افتد.
ارگانیسم های تک سلولی می توانند به صورت غیرجنسی از طریق تقسیم دوگانه، جوانه زدن ، یا هاگ سازی ، و از طریق جنسی توسط گامت ها یا از طریق کونژوگاسیون تکثیر شوند . 6
در یوکاریوت ها نسبت مساحت سطح به حجم کمتر از پروکاریوت ها است و بنابراین سرعت متابولیک کمتر و زمان تولید طولانی تری دارند.
مبدا
منشا سلول یوکاریوتی انقلابی ترین فرآیند بیولوژیکی از زمان پیدایش حیات از منظرهای مختلفی مانند مورفولوژی، تکامل تکاملی، ساختار ژنتیکی، روابط همزیستی و اکولوژی است. 7 تمام سلول های پیچیده از این نوع هستند و اساس تقریباً همه موجودات چند سلولی را تشکیل می دهند. 8 اگرچه توافقی در مورد زمان پیدایش یوکاریوت ها وجود ندارد، اما به طور کلی در اوایل پالئوپروتروزوییک حدود 2.5 میلیارد سال پیش پیشنهاد شده است. 9 10 تا کنون، قدیمیترین فسیلی که میتوان یوکاریوتی در نظر گرفت، 2.2 میلیارد سال سن دارد و به نام Diskagma شناخته میشود ، که نشاندهنده اولین نشانههای حیات چند سلولی همراه با موجودات زنده فرانسوی در 2.1 میلیارد سال پیش است. در جدایی بین یوکاریوت ها و گروه خواهر آنها باستانی آسگارد پایان دوره باستانی ( دوره نئوآرکائیک ) تخمین زده شد. 10

یوکاریا مرتبط هستند و ماشین آلات ژنتیکی با آرکیا از نظر DNA هسته ای و این دو گروه گاهی اوقات با هم در کلاد نئومورا طبقه بندی می شوند . از دیدگاه های دیگر، مانند ترکیب غشایی، آنها بیشتر شبیه باکتری ها هستند .
سه توضیح اصلی ممکن برای این پیشنهاد شده است: 12 13 14
- یوکاریوت ها از ادغام کامل دو یا چند سلول، سیتوپلاسم از یک باکتری و هسته از یک باستان به وجود می آیند.
- یوکاریوت ها از باستانی ها رشد کردند و ویژگی های باکتریایی خود را از پروتو-میتوکندری به دست آوردند.
- یوکاریوت ها و باستانی ها به طور مستقل از یک باکتری اصلاح شده رشد کردند.
شواهد فزاینده ای وجود دارد که به نظر می رسد نشان می دهد که منشا یوکاریوتی محصول ادغام یک باستان شناسی و یک باکتری است. در حالی که هسته سلول دارای عناصر ژنتیکی مرتبط با باستان است، میتوکندری و غشای سلولی دارای ویژگی های باکتریایی هستند. آمیختگی ژنتیکی زمانی آشکارتر می شود که تأیید شود که ژن های آموزنده به نظر منشا باستانی و ژن های عملیاتی با منشاء باکتریایی هستند. در هر صورت، این نیز درست است که تعداد معینی از صفات موجود منحصراً در یوکاریوتها با یک رویداد همجوشی به سختی توضیح داده میشوند. 15
فاصله میلیارد ساله بین منشأ یوکاریوت ها و تنوع آنها نیز مشخص نیست، زیرا باکتری ها بر زیست کره تا حدود 800 میلیون سال پیش 16 این فاصله پایداری محیطی، لیتوسفری و تکاملی به عنوان « میلیارد خسته کننده » شناخته می شود. 17 این تأخیر ممکن است صرفاً به دلیل دشواری ورود آن به زیست کره ای باشد که به طور کامل توسط پروکاریوت ها اشغال شده است. توضیحات دیگر مربوط به تکامل آهسته یوکاریوت ها یا افزایش اکسیژن است که تا پایان دوره مذکور به سطح ایده آل برای یوکاریوت ها نرسیده است. مناسبتر است طراحی بخشبندیشده سلول یوکاریوتی برای متابولیسم هوازی و معمولاً پذیرفته شده است که همه یوکاریوتهای موجود، از جمله بیهوازیها ، از اجداد هوازی دارای میتوکندری هستند. 18 19
تکامل

مدتی پس از ظهور اولین سلول یوکاریوتی، یک تشعشع انفجاری رخ داد که باعث شد آنها بیشتر سوله های اکولوژیکی موجود را اشغال کنند.
تکامل تک سلولی
اولین سلول یوکاریوتی احتمالاً تاژکدار بود، اگرچه دارای تمایلات آمیبوئیدی بود زیرا پوشش سفت و سختی نداشت. 21 از اجداد تاژک دار، برخی از گروه ها متعاقباً تاژک ها را از دست دادند، در حالی که برخی دیگر چند تاژک دار یا مژک دار شدند. گل مژه و تاژک که اسپرم را نگه می دارند ) ساختارهای همولوگ با 9 دوتایی میکروتوبولی سرچشمه می گیرند (از جمله آنهایی هستند که از سانتریول ها . 22
شخصیت آمیبوئید چندین بار در طول تکامل پروتیست ها پدید آمد و باعث پیدایش انواع شبه پاهای گروه های مختلف شد. این واقعیت که آمیبوئیدها از تاژکک ها می آیند و نه برعکس، همانطور که در گذشته تصور می شد، بر اساس مطالعات مولکولی (همجوشی، تقسیم یا تکثیر ژن ها، درج یا حذف اینترون ها و غیره) است. 23 24
به طور کلی پذیرفته شده است که کلروپلاست ها از درون همزیستی a سیانوباکتری ها و اینکه تمام جلبک های یوکاریوتی در نهایت از اجداد هتروتروف تکامل یافته اند . تصور میشود که تنوع سلولهای یوکاریوتی اولیه در میان تاژکهای جانوران رخ داده است: سلولهای شکارچی غیرفتوسنتزی با یک یا چند تاژک برای شنا، و اغلب برای تولید جریانهای آب برای شکار طعمه. 21
در حال حاضر در مورد محل قرار گرفتن ریشه درخت یوکاریا اختلاف نظر وجود دارد. پذیرفتهشدهترین احتمال، قرار دادن آن بین یا نزدیک به حفاریشده است ، که گروه پایه یوکاریوتها خواهد بود. 3 25
تکامل چند سلولی

در اولین بخش از تاریخ خود، یوکاریوت ها تک سلولی باقی ماندند. از دوره Ediacaran ، سلول های چند سلولی شروع به تکثیر می کنند، اگرچه این روند مطمئناً خیلی زودتر شروع شده است. موجودات تک سلولی زندگی استعماری شروع به انجام کارکردهای خاص در منطقه ای از مجموعه کردند. بدین ترتیب اولین بافت ها و اندام ها شکل گرفتند. چند سلولی به طور مستقل در گروه های مختلف یوکاریوت ها توسعه یافته است: گیاهان ، قارچ ها ، حیوانات ، جلبک های قرمز ، جلبک های قهوه ای و قالب های لجن . علیرغم چند سلولی بودن آنها، این دو گروه آخر هنوز در پادشاهی Protista طبقه بندی می شوند .
، جلبکهای سبز اولین گیاهان ، رشد کردند و اولین برگها را تشکیل دادند. در سیلورین اولین گیاهان زمینی و از آنها گیاهان آوندی یا کورموفیت ها پدید آمدند .
قارچهای تبدیل تک سلولی ردیفهایی از سلولها یا هیفها را تشکیل میدادند که در کنار هم، به موجودات چند سلولی جاذب با میسلیوم مشخص شدند . در ابتدا، قارچ ها آبزی بودند و اولین قارچ های خشکی احتمالاً در دوره سیلورین، درست پس از ظهور اولین گیاهان خشکی، ظاهر شدند. مطالعات مولکولی نشان می دهد که قارچ ها بیشتر با حیوانات مرتبط هستند تا گیاهان.
پادشاهی حیوانات با موجوداتی شبیه به پوریفرای مدرن که فاقد بافت واقعی هستند آغاز شد. بعداً آنها متنوع می شوند تا گروه های مختلف بی مهرگان و مهره داران را ایجاد کنند .
طبقه بندی و فیلوژنی


یوکاریوت ها Cavalier به طور سنتی به چهار پادشاهی تقسیم می شوند: Protista ، Plantae ، Animalia و Fungi (اگرچه -Smith 2004، 30 2015 31 جایگزین Protista با دو پادشاهی جدید Protozoa و Chromista شد ). این طبقه بندی دیدگاهی است که در حال حاضر پذیرفته شده است، اگرچه باید توجه داشت که پادشاهی Protista ، که به عنوان یوکاریوت هایی که در هیچ یک از سه گروه دیگر قرار نمی گیرند، تعریف می شود، پارافیلتیک است . به همین دلیل، تنوع پروتیست ها با تنوع اساسی یوکاریوت ها همزمان است.
طبقه بندی اخیر توسط Adl و همکاران. (2018) 3 از طبقهبندی به پادشاهیها اجتناب میکند، و آن را با طبقهبندی مطابق با فیلوژنی شناخته شده فعلی جایگزین میکند ، که در آن، از سوی دیگر، کلاسها یا گونهها نسبت داده نمیشوند دیگر به هیچ دستهای ، تا از ناراحتیهای ناشی از آن جلوگیری شود. برای طبقه بندی آنها به روز رسانی بعدی.
گروه های اصلی این طبقه بندی (معادل پادشاهی ها در طبقه بندی های قبلی) به شرح زیر است:
- Archaeplastida (= Primoplantae ). شامل گیاهان ، جلبک های سبز ، جلبک های قرمز و گلوکوفیت ها می شود . این موجودات فتوسنتزی کلروپلاست خود را با اندوسیمبیوز اولیه از یک سیانوباکتری به دست آوردند .
- سار ( S tramenopiles، Alveolata و R hizaria). این کلاد می تواند از نظر اجدادی با اندوسیمبیوز ثانویه جلبک قرمز فتوسنتز شود . Cavalier-Smith آن را در قلمرو Chromista گنجانده است .
- Stramenopiles (= Heterokonta ). این شامل جلبک های قهوه ای ، دیاتوم ها ، اوومیست ها و غیره است.
- آلوئولات . این شامل مژک داران ، آپی کمپلکس ها و داینوفلاژلات ها می باشد .
- ریزاریا . شامل روزن داران ، رادیولاریا و آمیبوفلاژله های مختلف است .
- آمیبوزوا . برخی از آمیب ها و قارچ های لجن .
- اپیستوکونتا . شامل حیوانات ، قارچ ها و choanozoans می شود .
- حفاری . مختلف تاژکهای هتروتروف یا فتوسنتزیکننده ، که دومی بهواسطه همزیستی ثانویه یک جلبک سبز است . این گروه احتمالاً پارافیلتیک هستند ، بنابراین باید به متامونادا ، دیسکوبا و مالاویمونادا تقسیم شوند .
برخی از گروههای پروتیستها دارای طبقهبندی مشکوک هستند، به ویژه کریپتیستا (کریپتوفیتها) و هاپتیستا (هاپتوفیتها)، در حالی که به نظر میرسد برخی دیگر خارج از گروههای بزرگ، به ویژه CRuMs ، Ancyromonadida و Hemimastigophora هستند .
علاوه بر این، دو گروه بزرگتر نیز شناسایی می شوند. Diaphoretickes (یا Corticata ) شامل Archaeplastida و Sar است ، در حالی که Amorphea (یا Podiata ) گروه Amoebozoa و Opisthokonta را شامل می شود . توجه داشته باشید که شکل آمیبوئید یا تاژک نشان دهنده تعلق به یک گروه طبقه بندی خاص نیست، همانطور که در طبقه بندی های سنتی اعتقاد بر این بود که گروه های مصنوعی را از نقطه نظر تکاملی ایجاد می کند (به polyphyly مراجعه کنید ).
زیر درخت فیلوژنتیک روابط بین گروه های اصلی یوکاریایی را بر اساس بورکی و همکاران نشان می دهد. 2019: 32
| یوکاریا [A] |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
افسانه ها: [A] هتروتروف تاژک دار اجدادی و یوکاریوت . [B] شیار تغذیه شکمی. [C] تاژک های ناهمگن . [D] آلوئول های قشری . [E] فیلوپودیا . [F] هاپتون و آکسوپودیا . [G] زائده روی تاژک قدامی. [H] ظاهر کلروپلاست ها با اندوسیمبیوز اولیه یک سیانوباکتری . ] همجوشی سهگانه ژنهای بیوسنتز پیریمیدین [ I . [J] Lobopodia . [K] تاژک عقب . [L] کیتین . [M] کلاژن ، بلاستولاسیون و بافت های تمایز یافته .